Alpha-Ketoglutarate: Benefits and Side Effects
;){kind=link}
;){kind=link}

Alpha-ketoglutarate (AKG) has been a popular sports supplement for a long time, but due to its roles in metabolism and aging, aging researchers have also taken an interest in it.
What is alpha-ketoglutarate?
AKG is a compound that is naturally produced in the body and is essential for energy production. It serves as a crucial intermediate in the metabolism of amino acids and is involved in various biochemical processes.
It was discovered in 1937 by Hans Adolf Krebs and William Arthur Johnson, who both worked at the University of Sheffield. Krebs later won the Nobel Prize for Physiology or Medicine in 1953.
The discovery of AKG was part of the overall identification of the citric acid cycle (the Krebs cycle), which is fundamental to cellular respiration and energy production in living organisms.
This cycle consists of a series of chemical reactions. It generates energy by oxidizing acetate, which comes from carbohydrates, fats, and proteins, and it produces carbon dioxide. It helps to both build muscle and to heal wounds.
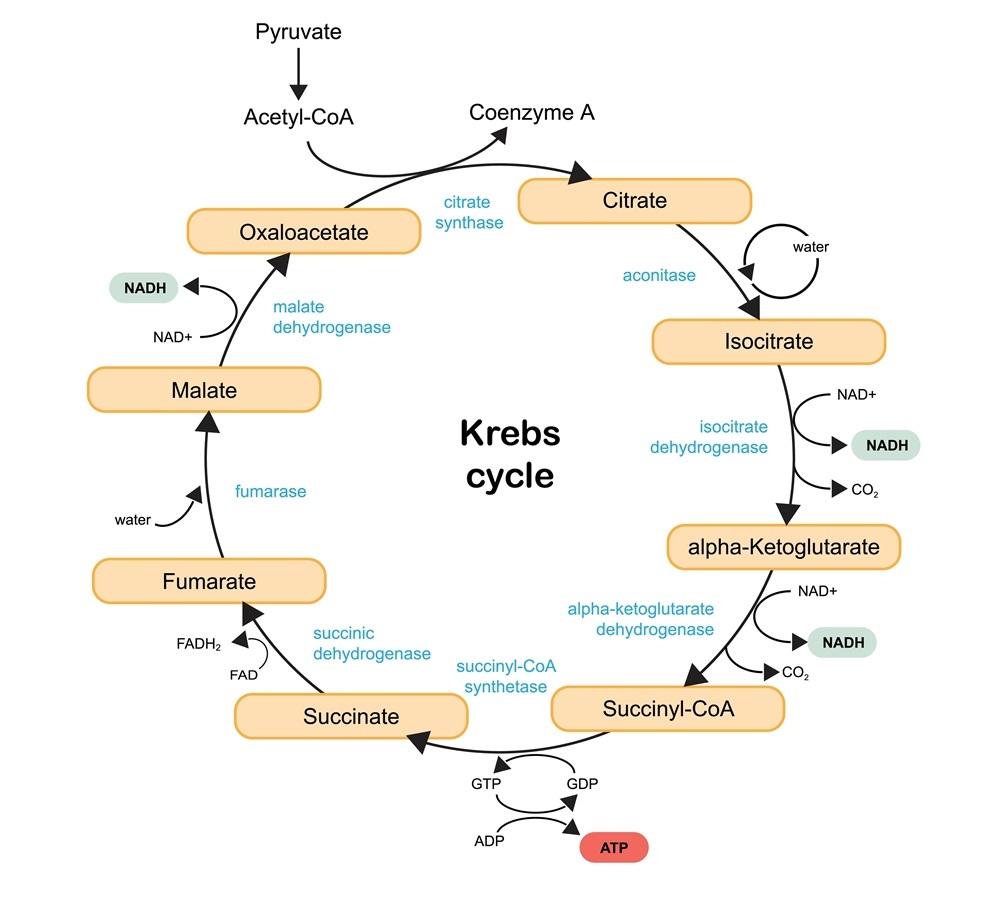
AKG is involved in a number of metabolic and cellular pathways. It acts as an energy donor, a precursor in amino acid production, and a cellular signaling molecule. It is also a regulator of epigenetic processes.
Sometimes, healthcare providers give AKG through an IV in order to help prevent damage to the heart from blood flow problems during heart surgery. It also helps prevent muscle loss after surgery or trauma.
AKG also acts as a nitrogen scavenger, allowing it to stop nitrogen overload and prevent too much ammonia from building up. It is also an important source of glutamate and glutamine, which build proteins and stop protein breakdown in the muscles.
Additionally, it regulates the ten-eleven translocation (TET) enzymes, which are involved in DNA demethylation. It also regulates the Jumonji C domain containing lysine demethylases, which are the major histone demethylases. In this way, it is an important player in gene regulation and expression.
The supplement industry also produces a synthetic version, which is chemically identical to naturally produced AKG. AKG supplements are popular among the sports community, especially with bodybuilders and athletes, as it is often said to boost endurance and lower fatigue during endurance training and is touted to help training improve both muscle growth and physical strength.
Can AKG slow down biological aging?
There is evidence that AKG can influence aging, and a number of studies suggest that this is the case. A 2014 study found that AKG can increase the lifespan of adult C. elegans by about 50%. It does this by blocking ATP synthase and the target of rapamycin (TOR) [1].
In this study, researchers found that AKG significantly increased lifespan and delayed some signs of aging. This included the loss of rapid coordinated body movement seen in older C. elegans.
ATP synthase
Mitochondrial ATP synthase is a ubiquitous enzyme involved in the energy metabolism of most living cells. ATP is a membrane-bound enzyme that acts as an energy carrier facilitating cellular energy metabolism. The 2014 study found that to increase lifespan in C. elegans, AKG requires ATP synthase subunit β and depends on the TOR pathway.
The researchers found that ATP synthase subunit β is a binding protein of AKG and discovered that AKG inhibits ATP synthase. This leads to a reduction of available ATP and decreased oxygen consumption along with an increase of autophagy in the cells of both C. elegans and mammals.
The direct binding of ATP-2 by AKG leads to the inhibition of enzymes. This also reduces ATP levels and oxygen consumption, increasing lifespan as a result. These effects are similar to when ATP synthase 2 (ATP-2) is genetically knocked out. From these findings, the researchers concluded that AKG likely increases lifespan by targeting ATP-2.
Essentially, what is happening here is that mitochondrial function is being partly suppressed, especially the electron transport chain, thus increasing lifespan in C. elegans. The key is to reduce mitochondrial function just enough without going too far and it becoming detrimental. The old saying “live fast, die young” is true. In this case, the worms live slowly and die old because of ATP suppression.
Alpha-ketoglutarate and TOR
TOR is part of a group of serine/threonine kinases from the phosphatidylinositol kinase-related kinase (PIKK) family. This pathway is conserved, which means it is found in many species, including C. elegans, mice, and humans. It regulates growth and metabolism and in mammals it is called the mammalian target of rapamycin (mTOR).
Various studies have found that blocking TOR can affect aging in different species. It can slow aging in yeast [2], C. elegans [3], fruit flies [4], and mice [5].
AKG does not directly interact with TOR, though it does influence it, mainly via the inhibition of ATP synthase. AKG depends at least partially on activated protein kinase (AMPK) and Forkhead box ‘Other’ (FoxO) proteins to influence longevity.
AMPK is a conserved cellular energy sensor found in multiple species, including humans. When the AMP/ATP ratio is too high, AMPK gets activated. This inhibits TOR signaling by activating the TOR suppressor TSC2 through phosphorylation. This process allows the cell to adjust its metabolism and balance its energy status effectively.
FoxO is a part of the Forkhead transcription factor family. It plays an important role in how insulin and growth factors affect many functions, including cell growth, metabolism, and cell death. One study found that to increase lifespan by reducing TOR signaling, the FoxO transcription factor PHA-4 is needed [6].
Alpha-ketoglutarate and autophagy
Finally, autophagy, which is activated by caloric restriction and the direct inhibition of TOR, is increased significantly in C. elegans given additional AKG. This means that AKG and TOR inhibition both increase lifespan. They may do this through the same pathway or through different pathways. Both pathways lead to the same final target.
Multiple studies have supported this idea. Research into starving yeast and bacteria has yielded similar results [7]. In humans after exercise, AKG levels are also higher [8].
This increase is thought to be a response to starvation. In this case, it involves gluconeogenesis, which activates certain enzymes in the liver. These enzymes create carbon from breaking down amino acids.
This matches the findings of the 2014 C. elegans study [1], which demonstrated that AKG levels rise in starving worms. However, AKG did not increase the lifespan of animals undergoing caloric restriction.
These results suggest that AKG is an important metabolite that helps control lifespan through starvation and calorie restriction. They also suggest that AKG connects cellular energy production and dietary restriction in lifespan regulation and that AKG could potentially slow down aging and treat age-related diseases.
Dr. Brian Kennedy published a mouse study on AKG demonstrating that it may help extend healthspan and lifespan [9]. Dr. Kennedy spoke about AKG at our EARD 2020 conference, where he explained how AKG may help treat aging and age-related diseases.
Alpha-ketoglutarate side effects
No significant side effects caused by AKG have been reported in humans, but if you do experience any adverse effects you should cease taking it immediately and consult your doctor.
Disclaimer
This article is only a very brief summary. It is not intended as an exhaustive guide and is based on the interpretation of research data. This article is not a substitute for consulting your physician about which supplements may or may not be right for you. We do not endorse supplement use or any product or supplement vendor.
Literature
[1] Chin, R. M., Fu, X., Pai, M. Y., Vergnes, L., Hwang, H., Deng, G., … & Hu, E. (2014). The metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR. Nature, 510(7505), 397-401.
[2] Kaeberlein, M., Burtner, C. R., & Kennedy, B. K. (2007). Recent developments in yeast aging. PLoS Genet, 3(5), e84.
[3] Hansen, M., Taubert, S., Crawford, D., Libina, N., Lee, S. J., & Kenyon, C. (2007). Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans. Aging cell, 6(1), 95-110.
[4] Luong, N., Davies, C. R., Wessells, R. J., Graham, S. M., King, M. T., Veech, R., … & Oldham, S. M. (2006). Activated FOXO-mediated insulin resistance is blocked by reduction of TOR activity. Cell metabolism, 4(2), 133-142.
[5] Selman, C., Tullet, J. M., Wieser, D., Irvine, E., Lingard, S. J., Choudhury, A. I., … & Woods, A. (2009). Ribosomal protein S6 kinase 1 signaling regulates mammalian life span. Science, 326(5949), 140-144.
[6] Sheaffer, K. L., Updike, D. L., & Mango, S. E. (2008). The Target of Rapamycin pathway antagonizes pha-4/FoxA to control development and aging. Current Biology, 18(18), 1355-1364.
[7] Brauer, M. J., Yuan, J., Bennett, B. D., Lu, W., Kimball, E., Botstein, D., & Rabinowitz, J. D. (2006). Conservation of the metabolomic response to starvation across two divergent microbes. Proceedings of the National Academy of Sciences, 103(51), 19302-19307.
[8] Brugnara, L., Vinaixa, M., Murillo, S., Samino, S., Rodriguez, M. A., Beltran, A., … & Novials, A. (2012). Metabolomics approach for analyzing the effects of exercise in subjects with type 1 diabetes mellitus. PloS one, 7(7), e40600.
[9] Shahmirzadi, A. A., Edgar, D., Liao, C. Y., Hsu, Y. M., Lucanic, M., Shahmirzadi, A. A., … & Kuehnemann, C. (2020). Alpha-ketoglutarate, an endogenous metabolite, extends lifespan and compresses morbidity in aging mice. Cell Metabolism, 32(3), 447-456.